
������ № 5
����� ����������������� – Polypodiophyta
������ ��������������� – Progymnospermae
����� ����������� – Ophioglossopsida
����� ������������ – Polypodiopsida
����� ����������������� – Polypodiophyta
����������������� ������� � ����� �������� ������� ����� ������ ��������. �� ����� ��������� ��� �������� ������ ���������� � ������������ � ����� �������������� ���� ������� � ������������. ������ � �� ��� ���� ����������������� ���������� ����������. � ��������� ����� ������������� ����� 300 ����� � ����� 10 ���. ����� ����������������� (���. 1).
���. 1. �������������������. 1 - ���������� ������������ (Athyrium filix-femina Roth.); 2 - ������� ������������ (Ophioglossum vulgatum L.); 3 - ���������� ���������� (Asplenium trichomanes L.); 4 - ��������� ������������ (Scolopendrium vulgare Sm.); 5 - �������� (Blechnum spicant With.); 6 - ����� ������������ (Pteridium aquilinum Kuhn).
����������������� �������������� ����� ������, ���������� �� ����� ������� ����, � ����������� � ����� ��������� �������������� – ������� � �������, ������ ��������, �������, �������� ������ � ������������ ������.
�� ���������� ������������ ����������� �� ������� ����������� �����. � ���������� �������������� � ����� ��������� �������� ����� � ����������������� ������������ ����� ������ ��������� ����� � �������� ����� ������� ������������ �� ������� �����, ���������� ��������, ��������������� ������������ � ��������. ��� � ����������� ������ ��������, �������� ����������, �������� ��������� – �������� – �������� �������������� ����� � �� ��������� �����.
����� � ���� ����������������� �������� �����������, ������ ���� ���������� – ���������� ������ ��� �������� �������� Ceratopteris.
��� � ����������� ������ ��������, �������� ����������������� ������� �������, �������������� ������ � ����� Hymenophyllaceae � � ���� Salvinia. ����� �����������, ������ �����������, � Ophioglossaceae – ����������� ����� ��������� � ����������. ��������� ������ ��������������.
������ ������� �� ������ ������ �������, ��� �������, ������ �� ����� � �������� ����������� ��� �������. ��� �� ����� ������ �������� ������������ �� �������� � ����������� ��������. ������������ ������� ����������� �����������������, ������� �� �������� ����� �������, �������� �������. ������� ������ ������ �������� � ��������� ��������������� ���������� �������, ���������� �� ������������. ����� ������� �������� ��� ��������, ��� �������� ����������, ��� ����� ���� ������� ��� ����� ��������, ������������, ����� ���������� ��� ���������������� ��������. ������ ������� �������� ������������� ��� ����� ��������� �����������.
��� ������������� ����� �������� �� ������ ����� � ������� ������, �� � �������� ��� �������� ��� �����, �������� ��� ������� � ���������. ������� ����� ���� �������������� ��� �����������, ����������� (��� Cheilanthes), �������� ����� �� ������� ������ (��� Blechnum). ������� ����� ���� ����������, �����������, ����� ����� �������� ����������, ���������� �����, ������� ����� ��������������� ��������. ��������� ����� ���������� (����������) � ���������� (����������). ���������� ����� ����������� ����� �� ����� ����� �����������, � �� ����� ���������. ������� ������ ������ ���������� ����� ��������, ������� ���������� ���������� ����.
������ ���������� ��������� ����������������� ���� ���������� �� ������� ����������, � �� ���������� ������� ������������ ����� ����������� ����������. � ��������� ����������� ����������������� ����������� ���������� – Schizaecaea – ���������, �������������� � � Gleicheiniaceae – ������������.
�� � ����������� ����������������� ���������� ������� ������������ ���������� ������� �����������. ��������� 2 ����: ����������� – ������ �������� ������� ������ �������, � ����������� – ������ �������� ������� � ����� ������, � �������� � � ����������.
����������� ����� ������������ ����� ������������ �������� ������� ���������� �����, �� ���� ���������� ������� ������������ ���� ���������� ������. ���������� ����� ���������� ������� ��������� ����������� ������. ����������� �������, ������������� � ����� ��� ������� ���������� �������� ������, �������� ��������� ���������, ��� ��������. ����������� �����������, � ������� ���������������� �������� ������� ����������� �������� ���� �� �����, �������� ������������ (�� ����. solen – «�����»), ���� �������� ������� � ����������� ����������� ����������� ����� ������, ��� ������ ����� ������ ������� ����������� ������� ����� �������, �� �������� ������������ (�� ����. diktyon – «����»). ����������� ������������ ����� �������� �������, ��������� �� ���� ��������������� ������. ��������� ����� ����������� �������� ���������� (�� ����. meros – «�����»). ����������� � ����������� �������� �������� ����������������� ������ ����� ����� ����������� ������������.
������ ������������, ����� ���������� ������, �� ������ ���������� ���������� �� ������� ����������� � �������� ������������. � �� ����� ��� �������� ������ ������������ ������������ ����� ������ ������� �� �������� ������ �������, � ������ ����������� �������� ��������������� �������� ���������, � ������������ ������ �������������� ������������� ����� ������� ������ �� ��������� ������� – ����������. ����� ���������� ��������������? ����������� ������������ ����������������� ����� ���������-������������ �������� ������, ������� ���� � �������� ����� ������� ��������. ����� ��������� � ����������� ������������ ���������� � �������� �����������. ��� ���� ������������ ���������� ����������� � ���������� ���� �������, ������ ������� ������� � ������ ����������� ����� �������� ��������� (���. 2).
���. 2. ����� �������� ����� �����������
1 – �������; 2 – ��������� �����; 3 – ���� ������� �������; 4 – ������� (��� ����� ������� �������); 5 – ������� �������; 6 – �����.
������� � ������� �������� ����� � ������� �������� �������� � ����� �������������� � ����������� �����. ������ ������������ ���������� ������� �������������. � ����������� ������� ������ ��������� ��� ������� – ���������� � ������������. � ������ (Matteuccia struthiopteris, Onoclea sensibilis, Drynaria) ������ ���������������� �� ���������� � ����������. ���������, ��� ��� 3 ���� ������� ������������ � ������ �������� ����������.
� ����������� �������������� ��������� Polypodiaceae ������� ����������� � ���������� ����������� ������� ���������� ��� ��� ���� (����������� �������� ������ ���������� � ��������� �����), ������� ���������� �������� ��������������� ���������. ��� ����������� ����� �������� ����� �������� ���������� ������� �������, � ��������� ����� ����������� ����� �� ���������� ����� (� ���������� �-�������� �����, � Plagiogyriaceae – Y-�������� �����).
������ �������� ����� ����� ���������� ������ – 1,2 ��� ���������. � Aspleniaceae ������ 3-7, �� � ����������� ������ 2, ����������� ������� � ����.
����������� ������ ����� �������������� ��������� (�����- ��� ���������������������). � ������������ ����������� ������ �������� – �������, ������ ��� �����������. ��������� ��������� ����� � ������� �� ��������������� ����� �������� ��� ����� (�� ����. rhachis – «�����������»), �������������� ����� ����������� �������. �������� ������������� ������� ����� ������ �����, ���������� �������� ���������� ������� (����� ���� ��������, ����������). � ������������ �������������� ���������� ����������� � ����������� ���� – ��������, �� ���������� ���� (������������� ��������� �������������� – Trichomanes reniforme). ��� ����� ����������� ���� ���������� �������� ����������, ������� �������� �� �����.
� ����������������� ����������� ����� ��� �������� ���� ���������� �������� (�� ����������� ������������) – �����������, ����������, ����������. ��� ���������� �������� ���������� � �������� ��������������� ���������.
����������� ������������ �������������� � �������� �� ���� ����. ����������� ����� – ������������� ��������. ��� ���� ������������ ���������� ���������� ������������������ ����������� ������� – ���������. � �������� ��������� ������������� � ������; � ����������� ������, ��������� ����� �����, �������� ��������. ���� � ����������� ����� ��������� ��������� ������������� �� ����� ������� ���� �� �������� ��������, ��� ���� ������ ��������� ������� ��������������� ������, �� � ����������� ������������ ��������� ��� ������ ������������� �� ������ ����������� �������. ����� ������������ ��������� � ����� ���������� ��������� (����). � ���� �������� ����������� ����������� ��������, � ��� ����������� ������������� ��� ���������� �����. ����� �����, ��������� �� �������� ������������� � ������������ ������������������, �������������, ���������� � ���������� ���������������. ���������� ����� ����� ����������� ���������� ���������� �� ��������. ��� ��� �������� – ����������� �������� � ���������� ����� ����� ��������� – ����������� ���������� � ������ ��������������� �������. ����������� ��������� �� �������� ����������, ���������� ������� �������� ��������������� ���������.
������ ������������ �������� � �������� �������� �� ������ ������������ �������. ��������, ��� ��������� ���������� ����������������� ����� ����������� (������������) ������������, �.�. ������ �� ��������� �������� ������� ������������� �������������� �����. ����� ���� ������� �������� ������� � ������������ ������������ ���������� ����������� � ����������� (Botrychium), ������������ � ����������� �����������. ��������� ��� ������������ ���������� – ������� (������������), ����������� � ������ ������������, ����� ������������� ��� ��������� ������������� �������� ���������.
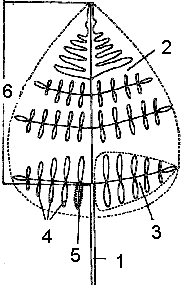
������������ ����� ������ � ����������������� �������� ������������ ��� ������� �� ������ ������� �������� ���������. ������ � ������������ ���������� �� ������ ������� ��������� ���������� � ������ ���������. ������� ��������� ������ ����������� �������� (� ��������� �� �������� �����). �� ��� � ��������� ������������ � ������� ������������� (Dicksonia, Loxsoma) ��� � ���� �������������� ��������� ������������� � ��������� �������������� �������, ��� ������ (�� ����. soros – «����»), ��� ���� ����������� ������������������, ����� ������������ ��������� ������������ ���������� (���. 3).
���. 3. ������������ ���������� � ������� ������������
1 – ��������� ������ (Pteridium); 2 – ������ ������ (Woodsia); 3 – ������ � ������������ ��������� (Dryopteris); 4 – ������, ������������� � ����� Asplenium, Athyrium, Polypodium.
� ������������� ��������� ����������� ����� ������� �������� � ���������� ���������� �� ����� ��� � ������ ��������� ������. � �������� �����������������, � ���� ������� �����������, �����������, ����������, ��������������, ���������, ������������ � ���������� ��� ��������� ����������� ������������ (������ ������), �.�. ����������� (������������� �������� ��������������, ������������ � ����������). ������, � ������� ���������� ������������� �������� � ���������� ����������, �������� ��������. �� ������������ ���������� – ��� ��������� ������� ������� ������������ ����� � ��������� ������������� � �������� ����.
����� – � ���������������� �������� � ���������� ���������� � ������. ��� ����������� ����������� ��������� �������� (�����������) ������ � ���������� ����� ���������� ��� ���������� �� ��������.
��������������� ������������� (�� ����. basis – «���������», �� ���. petere – «������������») ������������������, ����� �������� ���������� � ������� ����� �������� (� ���������, ������������� ��������� �������������� ����� �������, �������������� ����). ������ � ������������� ��������� �������� ���������� (�� ���. gradatio – «����������� ����������, ��������»). ����� ������ ����������� � ���������, �����������, � ���� �������������. ��������� ������ ��������� ���������� � ������ ������ ��������.
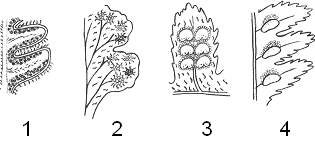
� ���������� ��������� ������ ������� – ��������� �������� ��������������. ���, � ���������� ������� ������ ������������ ������������ ����� �����, � ����� ������������������ ����� ����������� ������� (�� ���. indusium – «������� ������»). � ����������� �������� � �������� �������� ������� �����������.
������ ����� ������ – ��� ��������� � �������� (� ��������� �����������). ���������� – ��� Matonia, � �������� ��������� ��������� ����������� � �������� ��������.
������� ����� ����������� � ���������� ��������. �� ��������� ��������� � �������� ������ ��������� ����������� �������������� �� ����� - � ���������������.
� ��������������� ������������ ��������� ��������� �� ������ ������ � ����� ������������ ������, � ������������������ �� ��������� �� ����� ������ � ����� ����������� ������ (�� ����. leptos – «������»). ��� ����� �������� �������� ���������� � ���������� � ������� ������������. ��������� ���������� ���������� ����������� ������������. � ���������� ������ �� ������� ��������� ������� ��������� ������� ������ ���������� ������ – ����. ��� ���������� ��������� ���������� ������. ������ ��� � ������� ������������ �������� ��� ���������� ������ – ������� �� ������������� ������. ������ ����� �������� ������ ���������: ��������������, �����, ������������. ����������� ��������� �������� ������, ��� ���������, ����� ����������� – ��������, ��� ������� ����� ������ ������ �� �������� (��� ���������� �����); �� ���� ���������� ������ ������ (��������� ��������� �������� – Dryopteris filix-mas) (���. 4).
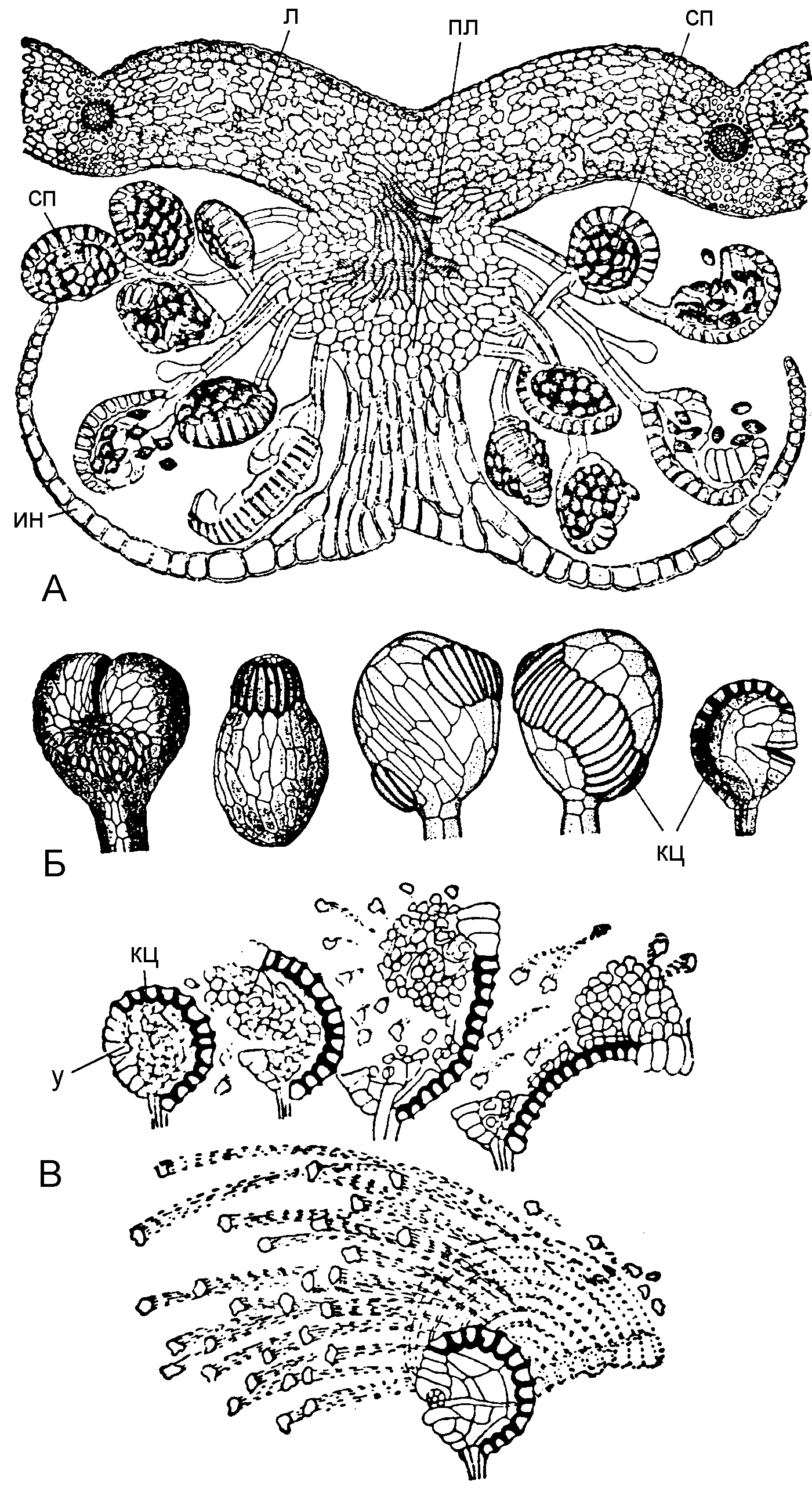
���. 4. �������� ������� � ���������� ������������
� – ���������� ������ ������ ��������� �������� (Dryopteris filix-mas); � – ������������ ����������; � – ����������� ����������; � – ����; �� – ��������; �� – ���������; �� – ������; � – �����; �� – �������.
� �������������� ����������� �������� ��������� �������, ����������������, �������� ������� ���������� ���� (8-15 ���.). � ����������� ���������� – �� 64 �� 16 ����, � ���� ������ ����������� ������������ ���������� � ���������� ���������� ����. ����� ��������� ����� (��������� ������ ��� ����) ����� ���������� � ��������� (��������).
���������� � �������� ����� ��������� ����� ����� � �������� ����������, �� � �����������, ���������, ������������ ���������� ����� ��������� �������������, ������ ����� �������� ��� ��������� �� ����������� �����. ������������ ���������� ������� �� ������������� ������� �����: ����� ���� ������������ �������� ��������, ������� – ������������ ����.
���� ����������� ��������� �������������� �� ���� ������� ������, � ����� �� ������� ������� ������������ ����������� ������, ������� ��������� ����������� ���������, ���������� ������������� ������ – ������������, ������������� ������������ ����������. ����������� ��� ��������� (�������, �������), ��� � ��������� ��������� ��������. �������������� ������������.
������������ ����������� ������ �������������� ����� ������������ ����������� ������� �� �������, �������, ������.
� ������ ������������� ������������ �������� ������ ��������: ������������� � ��������������� �������� ������������ ������� ���������, ����������� ������������, �������� ������������ � �������� ����������. ��������� ����� � ������������ (������������� � ���������������) �������� � ��������������� ���������. ����� ����������������� �������� ����� 300 ����� � ����� 10 ���. �����.
����� ������ � ���������� ���������� �����, ������ ����� ����������� ����������, ������� � ����������� ����� ������� � ����������� � ����� �������. ������ �� ��� �������� ����� � ���� ���� ������, ����� ��� ����� ������������, � ���������� �������; ������ ����� � ����� ���� ������������ �����. �� ������ ����������� ���������� �������, �������� � ��������, ������������������ ��������� ���� (�������). �� ������ ����������� ���������� ��������� ����������� ��������� � ����������� � ��� ����� ����������� ������ ������. �� ����� ������� �� ���������� ������ ������. ��������� ����� ��������� �� ������ ����������� ���������� � �������� ���������� ��������� �����.
� ��� ���������� ��������������, ��������� ����������� ��������������� �������������. ����� ��� ���������, �� ��� ����������� ���������� ��������� ��������, ����������� �������, ����������� � ����� ���������. � ��� ������ ���������� ��������������, � ��������� ������ ����� �� �������� ��������. ������� ������� ������ � ���������������� ��������������� �� �������� ��������, ��������� ����� ������� ������� �� ����������, �� ������ ������� ���������� ���������� ����������������� ��� ����������� ������ ������������ �������������. ����� ���������� �������� ��������� � ����� ��������� �����������.
��������� ����� ������ ��������������� – Progymnospermae, ������� ������������ �� �������� ������ �� �������� �������.
������ ������� �������� � ������������� �������� ������� �������� ������, � ����� ���������� �����. ����� �� ����������� �������� �������� ��������������� �� �����������������?
Ø ������� ������, �������������� ��������� �������;
Ø ������� �������� ������� � ������������ ������, ������� ������, ������������� �� ���������� � ������������� �������;
Ø ������������ ����������-����������� �������� ������, ����������� ��� ������������. �������� ���� ������ – ������� – ����� ����������� ��������, � ���������� – ������� – ����������� (��������) ��������.
� ����� ������������� ��������������� ������� ��������������� � ���������������, ��������� � ����� ������������. �������� �� ������������ ����������������� ������������� � ���� ������ ��������� ������������ ������������ ��������������, �������� ������������ �� � ������������. �������� ������� � ����������� �������, ��������� �� ������, - ����������������, ��� ������������� (Protopteridiales, ��� Aneurophytales), ����������� ��������������� � ������ �����������������. ���������� ��������������� ������������ � ������������ ����������� � �������������� ������� ��������������� (Archaeopteridales), ������ � �������� ������� �� �������� �������.
� ������ ��������������� �������������� ��� ������������ ���������: 1) ���������� �������� ��������, ���������������� ��������� ������������ ������ � ���������� ��������� �� ���� ���������; 2) ������� ������� ������� � ������������, � �� ��� – � ��������� �������; 3) ������������ ������������������ ����������� ������� – �������������� �, ��� ������� ������, �����������; 4) ���������� ���������� ���������� �� ��� �� ������; 5) ������� �� ��������������� � ���������������, ���������������� ����������� ���������� �������� � ����������� �� ��������; 6) ���������� ���������� �������������� �� ������������������; 7) ������������ ������ ��� ������������ ����, ������� �������������� ������ ��������������; 8) ������������ �� ������� ��������� ���������� ������, ������� ����������� �������� ����������� ����������� ���������.
������� ��� ��������������� ������
1. �� ������������ ������ ��������������� ���������� �����������: ��������������, �����������, ������������, ����������.
2. ����� �������-��������������� �������������� ������ ����������� (����������� �������� ����������, �������, ���������).
3. ��� ���������� ���������� ����������� �� ������� �������������?
4. ����� ����� �������������� ���������� ������������ � �����������.
5. ���������, � ������ ��������������� ������� ����������� �� �������-���������������� �������� (� �.�. �������� ������� ������������ � �����������)?
6. ����������, � ��� ����������� ���������������, �������� ����������� ������������ � �����������.
����� ����������� – Ophioglossopsida
����� ����������� ��� ������� ������������� ��� �������� ��������������� ���� ��� ���������� ���������������, ����� ������� ��������� ����� ��������, ������� ����������� �����������.
���������� � ������� ����� ������ �������������� �� ������� ����, ��� ���� ���� ������� ���� ����������� ������� ������ � �������� ��������� ����, � ����������� ����� ������� �������� �����������. ������������ ��� ���������������� ������� � ����������� ����� ���������� ��������� – �� ��� ����� � ����� �� �������, ����� ��������� � �������� ���������.
����������� – ��������� ��� ������� �������� �����������, ������ ������������ �����, �������� �� ������ � ������� ����� � ���������� ����� � �� �������� ������ (�����, �������, ������), �� ��������� ���� �������� �������� ���������. �� �������� ��������� ����, ����� ��� �����, ������� ������� �������� �����. � ������� ���� ����� ���������� ������ ���������� ����, ����������� � �����������. ��� ���������� ����� ������ �������� ��������.
����������� ������ – ����������� ������� �� ����������, �������� ������� � ������������ ������, ������� ������� ������������. ������ ��������� ����������� ��� ������ (�� 4-5 ���), ������� ������������� ��������. ������� �� ������� ����� ���� ������ ����������� ������������ ������� � ���������� ��������������� (�����������) ������������ � �������������, ���� � ������ ����������� ����� ����������� ����� ���������� � ����� �������������� ������� �������. ������ ����������� ���������� ���, ��� ��� �������� ��������� �� ��� ����� ������������� �� ����� � ������� ����� – ������������ (����������, �����������������) � ����������� (����������). ������������ �������� ����� ���� ����������� �������������, ����� ��������. ����������� �������� � ���� ���������� �������-�������������, �����������; � ���������������� ��� ����� ��������, �� � ������������ ������ � ������� ������������; � ���� ������� – �������, ��������.
��������� �������� ������� ��� ����������� ��������� �� ��������� �����, ���������, ��� ������. � ������� ��������� ����������� �������, ������������� �����. ����� ���������� ������ � �������, ��������� ����� ��������� ����� �����, �������� ����������, ������� �����������. ���������� ����������� ������������ �� �������� � ����� (���. 5). ��������� ����������� ������� ����������. �� ������ ���������� ������� ���������, ����� �������. ������� ����������� �� 1 ���� �� 10-20 ���. ����, � ����� � ������� ����������� ����������� ����� ����� �� ���� ������������, ����� � ������ �������� ��������.
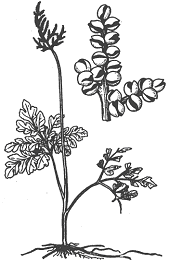
���. 5. �����������.
���������� (Botrychium lunaria) – ����� ���
� ������ ����������� ����������� ����������� � ������������ ������������� �����������. ����� ������ � ��� ���������� �� �����, ������������ �� ������. ��� ����������� – ���������� (������������) ���������, �� ������� ����������� �� �������� �������� � ������ �����. ��������������� � ��������� �������� �������� ������������� ����������, �� �������� ����������� ����� ������� � ������ ������������ � ��� ���������� �� ������������ ����� ����. � �������� ��������� ��������� ���� ������ ������������; � �������� ���� ���������� ���� ���� ������������ � «������������».
���� �������������� �������� ����������� �� �����, ��� ������������ � ������������� ������ ����������� ��������� ��������� � ����, � ������ ���� ��� ����� ��������� � ������. ���, ��� Botrychium simplex ������� � ������� ����� ��.
����, ��������� � ������� ����� ���������� ������� (������� ����� ���������� �������…, 2002):
��������� ����������� – Ophioglossaceae
���������� ���������� – Botrychium virginianum (L.) Sw.
�. ��������������� – B. multifidum (S.G. Gmel.) Rupr.
�. ������������� – B. lanceolatum (S.G. Gmel.) Angstr.
�. �������� – B. boreale Milde
������� ������������ – Ophioglossum vulgatum L.
����� ������������ – Polypodiopsida
����������� �����������, ������������� ������� ������� �������� � �������, ����������, ��������, ��������������� �� ���������������.
����������� ��� ����� ����� ���������� ��������, ����������� �� ������ ���� �� ����� �������. ������ ������������ ����� ������������ ������, � ����������� ���� ����������� ������ ������������ ������, ��� ���������� � ���������. ���������� ������� – �� ���������� �� ����� ������������������ ����� �����������. �������� ����������. ����� ����� ������.
������ ����� ��������� �����, ��� �� ��������, ��� � �� �����, ���������� � ���������. ������� ������ ��������� ���������.
������������� ��� ����� ������������� ��������. ��������� � ����������� ������� ������� � ������. ����������� ����� ���� ��� �������������. ��������� ����� ����������� ������ � ������ ������. ������� ������, ������� � �������� ������ ���������� ������� ������������� � ��������� ������� �������������, ��� ���� ����� ���� � ������ ��������� ���������� �����������. ���������� � ����������� �������������� �������, �������� ��� ���������, � ��������� ������� �� ����� ���������� ���������� � ���������� �����, �� � ������������� ���� ���������� �������������� � ������ ��������������.
������������� ��������� Polypodiidae � ������ ������������ 120 ������, ������������� 19 �������� � 46 �����. ��� �������� 2 ������������� ������: ������ (Dryopteris, Pteridium (���.6), Matteuccia � ��.) � ���������� ����������� (Cystopteris, Asplenium, Polypodium, Woodsia � ��.).

���. 6. ����������-����� (Pteridium aquiliunum)
� – ����� ���; � – ����� �������� � ��������; � – ����� ���������; � – ���������� ������ ��������� (�����); � – ����� ����� �� ���������� ������� (������� ����������): 1 – �������� ����, 2 – ���������� ����, 3 – ���������� ������������ �����, 4 – ���������� �����, 5 – ���������, 6 – ��������, 7 – ���������� ������, 8 – ������� ���������, 9 – ����������� ���������, 10 – ��������� �������, 11 - �������
�������� Salviniidae. ��� ������ ������������� �������� � ���������������, ������ ��������������� ������������, �������������� ������ �������� �����- � ���������.
������ ������������ ��� � ����������� ���. ����������� – ��� ������� �����������, � ������� ����������� ������ �����- � ���������������, � �����- � �������������� ���������� � ������ �������. ����� ���� ��������� – Salvinia – ������, �������. � ������ �������������� ��������� ��������� (���. 7). ������������� �������� ����� ��� ������ – Azolla, � ������� ����� �������� ����� ��������������� �������� �� ���� Anabena (���. Nostocaceae). ������ ���������� � ����������� ���������� �����.

���. 7. ��������� ��������� (Salvinia natans)
1 – ����� ���; 2 – ������� �������: �.� – ��������� ������; �.� – ��������� ����; �� – ������; 3 – ���������� ������ ����� �����- � ����������: �� – �������; � – �������; �� – ��������; ��� – ��������������; ��� – �������������; 4 – ������� ���������: � – �������; � – ������; �� – �������� ����������; �� – ��� �������������� ������; �.�� – ������ ������ ���������; ��� – �������������� ������; 5 – ������� ���������: �� – �������� ���������; �� –�������������� ������ ����������; 6 – ������� �� ����������; �� – ���������; � – ������.
����, ��������� � ������� ����� ���������� ������� (������� ����� ���������� �������…, 2002):
��������� ����������� – Aspleniaceae
�������� ��������� – Asplenium altajense (Kom.) Grub.
����������� ��������� – Camptosorus sibiricus Rupr.
��������� �������������� – Thelypteridaceae
���������� ������ – Oreopteris limbosperma (All.) Holub
���������� �������� – Thelypteris palustris Schott
��������� ������������ – Dryopteridaceae
����������� ����������� – Polystichum lonchitis (L.) Roth
�������� ������� – Dryopteris filix-mas (L.) Schott
����� �������, ����� �������� �������� ���������� ��� ������������ �����: ���������� – ��� ��������, ������� ����� �� ���� ������������ � ��������� ����� ������������� ���������, � ��� ��������� �������� ��������, ������� ����� �� ������� ���� – ���� ������������ � ��������� ����� ���������. �������� ������� ����������� �� ������, �� ������� �������� ������. ��������� ������������ ����� ��������� ��������.
��� ������������� �������� ��������, ��� �������� ������� ������� ������ – �������������, ����������� �������� ���� ��� ������� ����. � ������������� ������� ����� ����������� ���� ����� ���� �����, ����� ������ ��� ����� �������, �� ������� ���������� �� ��������������� ������, ������ ��� ���������� ������ ��������� ����, �������� �� �� ��� ��� ����������. �� ������ ������������� ������� ����������, ��������� ����� ����� ����, ������ ����������, �� ����� �������� ��������� ������ ��������, � ��� �������� ������ ���� ��������� �� ������������ ���� ��� ��������������. �������� ������� ������ � ����������� ������ ���� ����������� ��� ������� ����. ������ ���������� ����� ��������: ������������ � ���������������. ������� ��������� ������������.